| .. |
Scarabaeoidea of Southern South America Links
|
.
|
|
| |
| .. |
Scarab Guide Links
(goes to the University of Nebraska
web site) |
.
Classification |
|
|
| |
|
| . |
.
Hylamorpha elegans (Burmeister, 1844)
Aulacopalpus elegans Burmeister 1844:459. |
| |
|
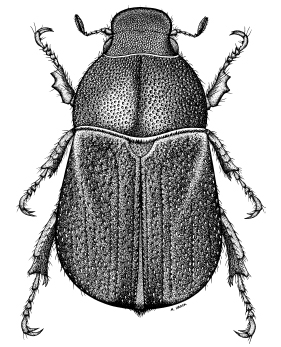
Dorsal habitus of Hylamorpha elegans (Burmeister). |
| |
|
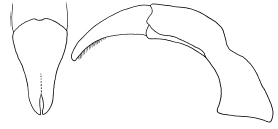
Figure 2. Male genitalia of H. elegans, caudal and lateral views.
|
| |
|
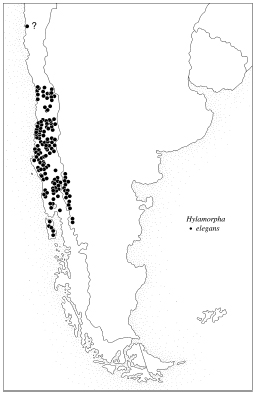
Locality records for H. elegans. |
| |
|
...... |
| Aulacopalpus elegans Burmeister 1844:459. NEOTYPE male at |
| |
UNSM (Lincoln),
labeled ‘‘Santiago, Chile, 1915’’// BCR and FCO red neotype label;
here designated to stabilize and fix the current concept of the name. |
| Callichloris perelegans Curtis 1845:449. LECTOTYPE at BMNH |
| |
(London),
labeled ‘‘type’’ (round label with red border// ‘‘Valparaiso’’ (hand-written)//‘‘Callichloris perelegans Curtis’’ (hand-written)// BCR lectotype
label; here designated to fix and stabilize the current concept of the
name. Synonymized by Solier 1851:91. |
| Aulacopalpus elegans var. australis Philippi 1861:741. |
| |
Unavailable name. |
| Sulcipalpus subviolaceus Nonfried 1894:125. Type not seen. |
| |
Synonymized by
Arrow 1901:401. |
| Hylamorpha cylindrica Arrow 1899:368. LECTOTYPE at BMNH |
| |
(London),
labeled ‘‘syntype’’ (round label with blue border)// ‘‘type’’ (round label
with red border)// ‘‘Chile’’ (hand-written on round label with ‘‘44/104’’
on reverse side//‘‘Hylamorpha cylindrica Arrow Type’’// BCR lectotype
label; here designated to fix and stabilize the current concept of the
name. NEW SYNONYMY. |
| Hylamorpha rufimana Arrow 1899:368. LECTOTYPE at BMNH |
| |
(London),
labeled ‘‘syntype’’ (round label with blue border// ‘‘type’’ (round label
with red border// ‘‘Chili 91–54’’// ‘‘Hylamorpha rufimana Arrow Type’’
(hand-written)// BCR lectotype label; here designated to fix and stabilize
the current concept of the name. Single paralectotype labeled ‘‘syntype’’
(round label with blue border// ‘‘Chili 91–54’’// BCR paralectotype. Synonymized
by Arrow 1901:400. |
|
| |
...... |
| |
Description. Length 11.8–18.2 mm; width across humeri 5.8–10.8 mm. Color of head
pronotum, and elytra light to dark apple green with metallic silver or bronze or orange
(rare) reflection usually present at apex of clypeus, mesad of each eye, on lateral margin
of pronotum, and on humeral and apical elytral umbones; occasionally color of head
and/or pronotum and/or elytra varies (in part or totally) to light brown or pale greenish
yellow; specimens preserved in fluid or killed in gases of certain chemicals may change
to cobalt blue, red, orange, or purple. Sternites and pygidium vary from light brown
(uncommon) to light or dark olive green (most common). Femora in males usually green
(similar to dorsum) or light brown (rare), females with profemora light brown and with
meso- and metafemora usually green (similar to dorsum); males with all tibiae green
and usually with strong, brassy reflections, especially on meso- and metatibiae; females
with protibiae light brown, meso- and metatibiae green and with weak to usually strong
brassy or bronzy reflections; tips of protibial teeth black. Thoracic sternites with dense, long, creamy white setae. Abdominal sternites and pygidium with creamy white, scalelike
setae; setae short, broad, moderately dense (surface visible between setae) to dense
(surface totally obscured); apex of pygidium at center with tuft of white setae. Head:
Surface entirely, densely rugopunctate (in smaller, darker specimens) or densely punctate
(in larger, lighter specimens); punctures moderately large to large, separated by less than
1 puncture diameter. Frontoclypeal suture finely impressed, arcuate. Clypeus with apex
broadly rounded, broadly reflexed. Interocular width equals 5.0-5.7 transverse eye diameters.
Antenna with 10 segments, club slender and slightly longer than segments 2-7 in male, subequal to segments 2-7 in female. Maxillary palpus with apical segment
lanceolate and with most of dorsal surface depressed. Mandibles subtriangular, outer
edge arcuate, apex acute to narrowly rounded. Pronotum: Surface densely punctate to
rugopunctate similar to that on head except punctures larger. Lateral edge with marginal
bead. Sides widest at about middle, strongly contracted from middle to acute apical angle.
Basal angle obtusely rounded. Base arcuate, without marginal bead, and with fringe of
dense, moderately long, white setae extending from beneath posterior margin. Scutellum
broadly subtriangular, apex narrowly rounded; surface densely punctate, punctures small.
Elytra: Surface wrinkled and with punctate striae; punctures small, setigerous; setae
short, broad. Intervals with sparse, small punctures, some (especially apically) with setae
like those of striae. Humeral and apical umbones well-developed. Lateral edge with
strong marginal bead. Epimeron broad below humerus, gradually narrowing posteriorly
for most of its length, and with sparse, long, white setae extending from just before
middle of elytron to anterolateral corners of pygidium. Pygidium: Surface densely and
finely rugopunctate but usually obscured by dense setae (often less so in females) and
with small tuft of longer setae at apex in center. Disc either side of middle slightly
depressed. In lateral view, surface weakly concave to almost flat. Legs: Protibia tridentate,
basal tooth slightly removed from others; tibiae wider in females. Protarsomeres 1-4 each with pair of small, straight spinules at apex on ventral side; segment 5 with small,
median tooth on ventral side in basal half, segment subequal in length to segments 1-4
in male, slightly shorter in female (male with first segment 2 times longer than second
segment, female with first segment 2.5-3.0 times longer than second segment). Male
with one claw larger than other, female with claws subequal in size; apices of all claws
simple. Meso- and metatibiae each with short, obliquely transverse carina on external
edge just past middle; apices of tibiae weakly expanded and with row of 12-18 small
spinules. Venter: Mesometasternal process absent. Sternites 5-6 longer than 1-4, last
sternite with apex arcuate and entire in both sexes. Thoracic sternites and posterior edges
of femora with moderately dense, long setae; abdominal sternites with dense, elongated,
scale-like setae. Parameres: Figure 2. |
| ...... |
|
| |
Neotype. Inasmuch as there has been considerable confusion about the correct
identity of the several ‘‘species’’ included in Hylamorpha (as well as the
status of the genus itself, i.e., Hylamorpha, Callichloris, Sulcipalpus, or Aulacopalpus),
we believe it is essential to fix the type specimen for H. elegans.
Burmeister (1844) did not designate a type for A. elegans. A type specimen,
if such ever existed, was not found by BCR in searches of the Burmeister
collection at Martin Luther Universität in Halle, the large collection of the
Zoologische Staatssammlung in Munich, the historical collection of the Deutsche
Entomologisches Institut in Eberswalde, or the principal collection in Germany
at the Museum für Naturkunde in Berlin. In fact, we believe that Burmeister
never designated a "typus" because he was referring to the previously
existing name of Callichloris elegans in Dejean's catalog (the name is unavailable/
nomen nudum because it was never accompanied by a description or
reference to one). Burmeister did not believe he was creating a new species
but simply referring to a known species listed in a catalog. He became the
author of elegans because he was the first to actually describe it. Accordingly,
we designate a neotype here.
Burmeister (1844) indicated his specimen was from near Valparaíso, Chile. However, we have been unable to locate unbroken male specimens with a
label that indicates Valparaíso. Not wanting the neotype to be broken (hence,
incomplete), we chose another specimen that agrees with Burmeister’s original
description and is geographically close to Valparaíso. |
| |
|
|
Distribution. Hylamorpha elegans is found on both sides of the Andes in
central and southern Chile and southwestern Argentina. Its distribution coincides
remarkably with the distribution of Nothofagus species (Nothofagaceae)
on which it is known to feed. |
| |
|
| |
Locality Records (see map). 1,930 specimens examined.
ARGENTINA (663). CHUBUT (84): Epuyén, Lago Puelo (6 km N); NEUQUEN (30): Isla Victoria, Lago Lacar, Parq. Nac. Lanín, Río Quilquihue (San
Martín de los Andes), San Martín de los Andes (9 km NE); RIO NEGRO
(549): Bariloche, El Bolsón, Río Villegas (ruta 258).
CHILE (1267). ACONCAGUA (1): No data; ARAUCO (72): Caramávida,
Caramávida (20 km W), Contulmo, Pichinahuel; Biobío (131): Bullileo, El
Abanico, El Quilleco (30 km E), Santa Bárbara (25 km E); CAUTIN (106):
Fundo Chacamo (35 km NW Nueva Imperial), Fundo El Coigue (29 km NW
Villarrica), Fundo Las Selvas (750 m NW Nueva Imperial), Loncoche, Loncoche
(12.3 km N), Malalcahuello, Nueva Imperial (750 m NW), Pucón, Río
Trancura (Villarica), Temuco, Temuco (20 km E), Villarrica, Villa Portales (7
km W); CHILOE (28): Ancud, Castro, Chiloé Island (30 km S Ancud), Hueque
Trumao (Chiloé Island, 22 km N Quellón), Lago Tepuhueco (40 air km SW
Castro); CONCEPCION (47): Fundo Andalién, Fundo Pinares, Hualpencillo,
Lirquén, Penco, Punta Hualpén, San Rosendo, Santa Juana; COLCAGUA (2):
Peumo; COQUIMBO (2): Ovalle, Parq. Nac. Fray Jorge; CURICO (104): El
Coigo, Fundo La Montana (6 km E Los Queñes), Las Trancas, Los Niches;
LINARES (25): Parral, Tranque de Bullileo; LLANQUIHUE (15): Departamento
de Maullín, Ensenada, Lago Chapo; MALLECO (255): Angol, Angol
(6 km W), Chanchuco, Cordillera Nahuelbuta, Cordón Las Raíces, Curacautín,
Parq. Nac. Nahuelbuta, Perquenco (10 mi. N), Pillín Pilli, Pino Hachado, Termas
de Manzanar, Termas de Río Blanco, Vegas Blancas (27 km W Angol),
Victoria, Victoria (4 km W), Villa Portales; MAULE (183): Cauquenes (400
m W), Cayurranquil (400 m W Cauquenes), Chovellén, Constitución, El Pantanillo
(17 km SE Constitución), Forel, Paso García (23 km NW Cauquenes),
Pailahueque, Pelluhue, Río Teno (Teno), Tregualemu; ÑUBLE (79): Alto Tregualemu
(20 km SE Chovellén), Cobquecura, Cueva de los Pincheira (Recinto),
Curanipe (17.5 km S), Las Trancas (21 km E Recinto), San Carlos (18
km E), San Carlos (40 km E), San Carlos (50 km E), Recinto, Recinto (4 km
SE), Recinto (10 km W), Recinto (13 km E); OSORNO (7): Cunco, Osorno,
Puyehue, Puyehue (10 km E); SANTIAGO (127): Aculeo, Bucalemu, El Canelo,
El Portezuelo (7 km N Santiago), La Herrera, Las Condes, Llolleo, Maipú, Punta Yeso (70 km SE Santiago), Río Blanco, Río Colorado (40 km SE
Santiago), Río Peuco (Pilay), Santiago, Santiago (cuesta La Dormida); TALCA
(18): Fundo El Radal, Talca (22 mi N), Vilches, Vilches (5 km W), Vilches
Alto; VALDIVIA (27): Panguipulli, Pucará, Río Licán (N side Lago Puyehue),
Santo Domingo; VALPARAISO (16): Algarrobo, Isla Teja, Limache, Valparaíso. |
| |
|
| |
Temporal Distribution. January (217), February (97), March (14), April
(3), May (33), September (261), October (13), November (219), December
(852). Gutiérrez (1949) reported that H. elegans was one of the most common
species of scarabs in the Cordillera de Pemehue in a June 1946 expedition, and that hundreds of dead specimens were found on the ground in December
and January. We saw no museum specimens from June to August. |
| |
|
| |
Remarks. Various catalogs (e.g., Ohaus 1918; Blackwelder 1944; Machatschke
1965, 1972) have already included the synonymy of all but H. cylindrica under H. elegans. We concur with these synonymies based upon examination
of the types and careful analysis of the original descriptions. Hylamorpha
cylindrica (a morphotype that is slightly larger and lighter in color)
is conspecific with H. elegans and is synonymized here.
Philippi (1861) described Hylamorpha elegans var. australis and explicitly
referred to it as a "climatic variety" of H. elegans. Accordingly, this name is
unavailable because it referred to an infrasubspecific entity described as a
variety before 1961 (International Commission on Zoological Nomenclature
1999; Article 45.6.4).
Hylamorpha elegans is variable in size, color, surface sculpturing, amount
of setae, and even body shape (somewhat flattened to highly vaulted). Ohaus
(1905) correctly noted that darker specimens tended to have more strongly
punctate, wrinkled, and setose elytra. Sexual dimorphism is distinct. Males
nearly always have green protibia (even if the meso- and metatibiae are brown)
whereas females have brown protibiae; males have the first segment of the
protarsus a little shorter than in females; males have one claw on all legs larger
than the other claw whereas all the claws are subequal in size in females; and
males have the antennal club a little longer than in females.
Hylamorpha elegans is often locally abundant. Although frequently associated
with Nothofagus forests, it is also found feeding on other native plants
and trees and is sometimes an agricultural pest (Pedro Vidal, pers. comm. to
BCR, 2000). It occasionally causes severe defoliation of Nothofagus species
[principally N. antarctica (Forst.), N. betuloides (Mirb.), N. dombeyi (Mirb.),
and N. obliqua (Mirb.)], and young trees have even been known to be killed
by huge numbers of H. elegans feeding on them.
(Carrillo and Cerda 1987; Gentili and Gentili 1988; and Veblen et al. 1996).
Glare et al. (1993) reported that the fungus, Beauveria vermiconia (de Hoog
and Rao) (Deuteromycotina), was a pathogen of H. elegans in Osorno, Chile;
in their observations, 20–30% of a population of late third instar larvae were infected.
During January at Lago Traful (Argentina, Neuquén Province) it is very
common to see rainbow trout [Oncorhynchus mykiss (Walbaum)] feeding on
adults of H. elegans that fall into the lake water from Nothophagus trees.
Hylamorpha elegans is exceedingly abundant on the shores of this lake (and
presumably others), and it is a common occurrence to catch trout with their
stomachs full of these scarabs (FCO, pers. obs.).
Cartagena (1975) tested success of egg-hatch in different soil types, and
found that soils high in sand content favored increased eclosion.
In Chile, H. elegans, along with species of Brachysternus, are referred to
as "San Juanes" ("San Juan" for a single specimen); the origin of the name
is unknown (P. Vidal, pers. comm., March 2000). They are also called "pololos
verdes" (Arias 2000) or also simply "pololos". |
| |
|
| |
Excerpt from:
Ratcliffe, B.C. and F.C. Ocampo . 2002. A review of the genus Hylamorpha (Coleoptera: Scarabaeidae: Rutelinae: Anoplognathini, Brachysternina). The Coleopterists Bulletin, 56(3):367–378. |
| |
|