 |
|||||||
|
|||||||
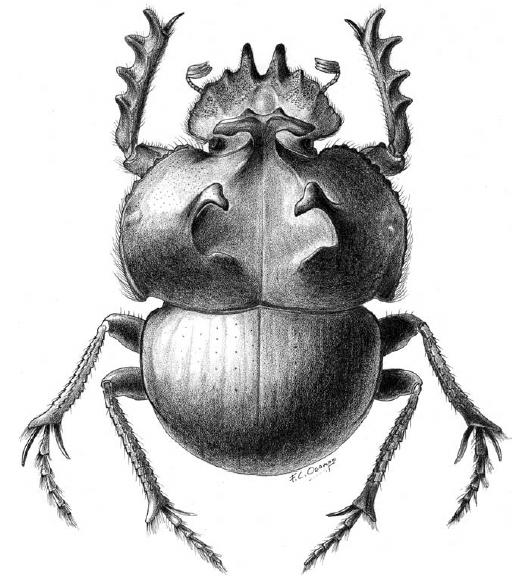
Glyphoderus monticola Burmeister.
Illustration by Federico Ocampo.Distribution of Glyphoderus species
.
|
||
| Scarabaeinae Taxa Map | ||
| Scarabaeinae
Key |
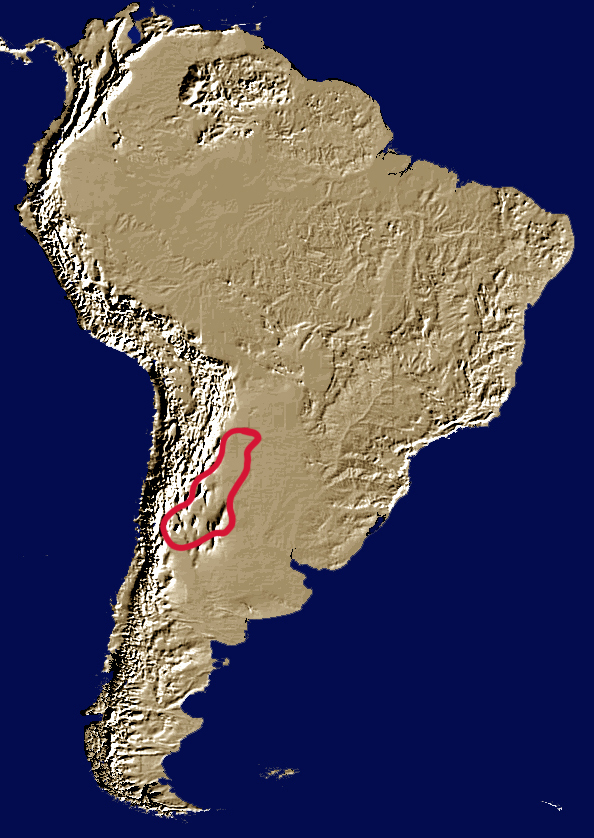
Diversity and distribution: 3 species. The genus Glyphoderus is restricted the Monte and southeastern Chacoan biogeographic provinces in northwestern Argentina, between 150–1,850 m altitude.
Revision or Synopsis: Ocampo, F. C. 2004. Food relocation behavior and synopsis of the southern South American genus Glyphoderus Westwood (Scarabaeidae: Scarabaeinae: Eucraniini). The Coleopterists Bulletin 58: 295-305.
Figure 2. Female of Glyphoderus sterquilinus (Westwood) carrying a rodent dung pellet.
Photo by Federico Ocampo.

Biology: Under natural conditions, the period of surface activity of G. monticola and G. sterquilinus is from 9:30 am to 1:00 pm, and from 5:00 pm to 7:00 pm, depending on atmospheric and soil temperature. No nocturnal, surface activity was recorded. During the observations atmospheric temperature varied between 21–40 408C in the shade. Both species prefer open sandy or clay soils with a 20–60% vegetation cover. Glyphoderus sterquilinus and G. monticola monticola, as do most of the Eucraniini, specialize on dehydrated dung pellets. Specimens were observed carrying pellets of ‘‘vizcacha’’ (Lagostomus maximus (Desmarest)), ‘‘cuis chico’’ (Microcavia australis (Geoffroy Saint-Hilaire & D’ Orbigny)), and goat. Specimens of G. sterquilinus did not take dry horse or cow dung even when it was offered, and they were not attracted to dung traps baited with fresh human or cow dung. When foraging, they run on four legs, keeping their forelegs motionless and in a horizontal position with respect to the surface. Adults run in zig-zags or do not follow any particular direction from their burrow (apparently searching randomly). To carry food, the beetles grasp it with the foretibiae and run forward using only their middle and hind legs (Fig. 2), this behavior is known only for members of the tribe Eucraniini. It is not clear how the beetles find their way back to the burrow. As described by Byrne et al. (2003) and Dacke and Warrant (2002) for the South African flightless dung beetles Scarabaeus rugosus (Hausman), S. rusticus (Boheman), Kheper nigroaeneus (Boheman) and Pachylomerus femoralis (Kirby) (Scarabaeinae: Scarabaeini) Glyphoderus species must rely on cues such as the pattern of polarized light. Beetles may use polarized light as a compass bearing that can be used to ‘‘calculate’’ the direction to the burrow. When the entrance of the burrow is reached, the beetles enter by walking forward rather than backward. Observations published by Zunino et al. (1989) regarding the above behavioral character, suggested that the beetles drop the dung pellet turn back, and enter the nest walking backward dragging the pellet. I have not observed this for any Eucraniini species. The burrow is always previously dug. It is variable in depth (usually between 0.25–1 m long) and variable in slope (between 35–60 608 with respect to ground surface). The tunnel may be straight or curved. The depth of the burrow is apparently dictated by the level of soil moisture. Burrows are sometimes bifurcated near the end. The beetles store the food at the end of the burrow and no special chamber seems to be prepared for that purpose. Usually between 3–10 dung pellets are stored, and each pellet is carried independently. Sexual cooperation has been observed and it follows the same behavior as that observed for individuals. No special brood chambers were observed in burrows constructed by pairs. During the hours where there is no surface activity, it is common to observe the entrance of the burrow obscured with sand or the available substrate. The behavior of G. centralis is similar and it was described by Zunino and collaborators in 1989.
Species profiles: (click on species names)
Glyphoderus centralis
Glyphoderus monticola
Glyphoderus sterquilinus
Literature cited
|
||||
|
|